Большинство растений усваивает неорганический углерод именно по пути цикла Кальвина. Однако довольно большая группа растений (около 500 видов) тропического происхождения выработала в процессе эволюции некоторую модификацию процесса, усваивая неорганический углерод путем образования в результате его акцепции четырехуглеродных соединений. Это растения, приспособившиеся к фотосинтезу в условиях повышенной температуры воздуха и избыточной освещенности, а также пониженной влажности почвы (засухи). Из культурных растений обладают таким метаболитическим процессом кукуруза, просо, сорго, сахарный тростник. У ряда сорных растений также наблюдается именно эта особенность метаболизма (свинорой, просо куриное, щирица) и т.д.
Особенностью анатомического строения таких растений является наличие фотосинтезирующих клеток двух типов, которые располагаются в виде концентрических кругов - радиально расположенные вокруг проводящих пучков клетки обкладочной паренхимы и мезофилла. Этот тип анатомического строения называется кранц-типом (от немецкого Кranz -венок).
Этот тип метаболизма был изучен в 60-е годы прошлого века, большую роль сыграли при этом исследования советских ученых Карпилова, Незговоровой, Тарчевского, а также австралийских ученых Хэтча и Слэка. Именно они предложили законченную схему цикла, поэтому принято этот процесс называть также циклом Хэтча-Слэка-Карпилова.
Процесс происходит в два этапа: поступающий в мезофилл СО 2 вступает в соединение с трехуглеродным соединением (ФЕП) - фосфоенолпировиноградной кислотой - которая превращается в четырехуглеродное соединение. Это и есть ключевой момент процесса, из-за которого он и получил свое название, так как неорганический углерод, акцептируясь трехуглеродным соединением, превращается в четырехуглеродное соединение. В зависимости от того в какое именно четырехуглеродное соединение превращается неорганический углерод различают три группы растений:
НАДФ-МДГ образуют яблочную кислоту при участии фермента малатдегидрогеназы, а затем пировиноградной кислоты,
НАД-МДГ образуют аспарагиновую кислоту и аланин,
ФЕП-КК образуют аспарагиновую кислоту и фосфоенолпировиноградную кислоту.
Наиболее значимые для сельского хозяйства растения относятся к НАДФ-МДГ типу.
После образования четырехуглеродного соединения происходит его перемещение во внутренние клетки обкладочной паренхимы и расщепление или декарбоксилирование этой молекулы. Отделившаяся карбоксильная группа в виде CОО - входит в цикл Кальвина, а оставшаяся трехуглеродная молекула - ФЕП - возвращается опять в клетки мезофилла.
Такой путь фиксации углекислого газа позволяет растениям накапливать в виде органических кислот запас углерода, проводить процесс фотосинтеза в наиболее жаркое время дня при сокращении потерь воды на транспирацию за счет закрытия устьиц. Эффективность использования воды такими растениями в два раза больше, чем у растений, происходящих из умеренных широт.
Для С 4 -растений характерны отсутствие обратного потока углекислого газа при фотодыхании и повышенный уровень синтеза и накопления органических веществ.
Поскольку в цикле Кальвина первичными продуктами включения неорганического углерода в органический являются трехуглеродные соединения, данный процесс носит название С-3 путь фотосинтеза.
Важно заметить, что для синтеза одной молекулы глюкозы должно произойти шесть оборотов цикла Кальвина. В каждом обороте используются три молекулы АТФ (две для активирования двух молекул фосфоглицериновой кислоты и одна при регенерации рибулезодифосфата) и две молекулы НАДФ. Н 2 для восстановления кислоты в альдегид. Таким образом для синтеза одной молекулы глюкозы крайне важно потратить 12 молекул НАДФ. Н 2 и 18 молекул АТФ.
Важно отметить, что физиологическое значение цикла Кальвина состоит не только в акцепции углекислого газа, но и в создании массы углеводных соединений, которые идут как на синтез запасных веществ, так и на создание компонентов хлоропласта и текущий метаболизм клетки. .
Большинство растений усваивает неорганический углерод именно по пути цикла Кальвина. При этом довольно большая группа растений (около 500 видов) тропического происхождения выработала в процессе эволюции некоторую модификацию процесса, усваивая неорганический углерод путем образования в результате его акцепции четырехуглеродных соединений. Это растения, приспособившиеся к фотосинтезу в условиях повышенной температуры воздуха и избыточной освещенности, а также пониженной влажности почвы (засухи). Из культурных растений обладают таким метаболитическим процессом кукуруза, просо, сорго, сахарный тростник. У ряда сорных растений также наблюдается именно эта особенность метаболизма (свинорой, просо куриное, щирица) и т.д.
Особенностью анатомического строения таких растений является наличие фотосинтезирующих клеток двух типов, которые располагаются в виде концентрических кругов - радиально расположенные вокруг проводящих пучков клетки обкладочной паренхимы и мезофилла. Этот тип анатомического строения принято называть кранц-типом (от немецкого Кranz -венок).
Этот тип метаболизма был изучен в 60-е годы прошлого века, большую роль сыграли при этом исследования советских ученых Карпилова, Незговоровой, Тарчевского, а также австралийских ученых Хэтча и Слэка. Именно они предложили законченную схему цикла, в связи с этим принято данный процесс называть также циклом Хэтча-Слэка-Карпилова.
Процесс происходит в два этапа: поступающий в мезофилл СО 2 вступает в соединение с трехуглеродным соединением (ФЕП) - фосфоенолпировиноградной кислотой - которая превращается в четырехуглеродное соединение. Это и есть ключевой момент процесса, из-за которого он и получил свое название, так как неорганический углерод, акцептируясь трехуглеродным соединением, превращается в четырехуглеродное соединение. Учитывая зависимость оттого в какое именно четырехуглеродное соединение превращается неорганический углерод различают три группы растений:
НАДФ-МДГ образуют яблочную кислоту при участии фермента малатдегидрогеназы, а затем пировиноградной кислоты,
НАД-МДГ образуют аспарагиновую кислоту и аланин,
ФЕП-КК образуют аспарагиновую кислоту и фосфоенолпировиноградную кислоту.
Наиболее значимые для сельского хозяйства растения относятся к НАДФ-МДГ типу.
После образования четырехуглеродного соединения происходит его перемещение во внутренние клетки обкладочной паренхимы и расщепление или декарбоксилирование этой молекулы. Отделившаяся карбоксильная группа в виде CОО - входит в цикл Кальвина, а оставшаяся трехуглеродная молекула - ФЕП - возвращается опять в клетки мезофилла.
Такой путь фиксации углекислого газа позволяет растениям накапливать в виде органических кислот запас углерода, проводить процесс фотосинтеза в наиболее жаркое время дня при сокращении потерь воды на транспирацию за счёт закрытия устьиц. Эффективность использования воды такими растениями в два раза больше, чем у растений, происходящих из умеренных широт.
Для С 4 -растений характерны отсутствие обратного потока углекислого газа при фотодыхании и повышенный уровень синтеза и накопления органических веществ.
Фотосинтез – это процесс трансформации поглощенной растением энергии солнечного света в химическую энергию органических соединений. С 4 - путь фотосинтеза или цикл Хетча-Слэка.
Австралийскими учеными Хетчем и Слэком был описан С 4 -путь фотосинтеза, характерный для тропических и субтропических растений (сахарный тростник, кукуруза и др.). Листья этих растений содержат хлоропласты двух типов: обычные в клетках мезофилла и крупные хлоропласты, не имеющие гран и фотосистемы II, в клетках обкладки, окружающих проводящие пучки.
В цитоплазме клеток мезофилла присоединяет СО 2 к пировиноградной кислоте, образуя щавелевоуксусную кислоту. Она транспортируется в хлоропласты, где восстанавливается до яблочной кислоты при участии НАДФН. В присутствии ионов аммония щавелевоуксусная кислота превращается в аспарагиновую кислоту. Яблочная и (или) аспарагиновая кислоты переходят в хлоропласты клеток обкладки, восстанавливаются до пировиноградной кислоты и СО 2 . СО 2 включается в цикл Кальвина, а пировиноградная кислота переносится в клетки мезофилла, где превращается в пировиноградную кислоту. Такой механизм позволяет растениям фотосинтезировать при закрытых из-за высокой температуры устьицах. Кроме того, продукты цикла Кальвина образуются в хлоропластах клеток обкладки, окружающих проводящие пучки. Это способствует быстрому оттоку фотоассимилятов и тем самым повышает интенсивность фотосинтеза.
Липиды: классификация и их роль в растении.
Липиды представляют собой достаточно сложные по химической структуре вещества. В их состав также входят углерод, кислород, водород, но в отдельные группы липидов могут входить и фосфор, и сера, и азот (фосфатиды, пигменты). Все липиды гидрофобны, т.е. не растворяются в воде. Функции у липидов различны в зависимости от химического строения. Липиды не являются биополимерами.
Липиды классифицируются на 5 больших групп по признаку функции и сложности строения:Жиры- наиболее легко синтезируемая группа липидов. С химической точки зрения - это эфиры жирных кислот и глицерина.Основные функции жиров - энергетическая, строительная и запасающая.Воска - это жироподобные вещества, твердые при комнатной температуре. По химической структуре - это сложные эфиры между жирными кислотами и высокомолекулярными одноатомными спиртами жирного ряда.Основная функция восков - защитная.Фосфатиды - к которым относятся глицерофосфатиды, лецитины и кефалины - это молекулы сложных эфиров глицерина, жирных кислот и фосфорной кислоты. Эти вещества входят в состав запасных жиров и предохраняют их от прогоркания.Основная функция фосфатидов - запасающая.
Пигменты (хлорофиллы и каротиноиды) - это особая группа липидов, имеющая сложное строение, куда входят и азотистые радикалы. К пигментам относят две группы веществ - хлорофиллы и каротиноиды.
Основная функция пигментов - участие в энергетической (световой) фазе фотосинтеза.Стероиды - это производные сложного гетероциклического соединения. В эту группу соединений входят высокомолекулярные спирты (стеролы) и их сложные эфиры (стериды) Наиболее известный стероид - эргостерол, из которого в промышленности получают витамин Д.
Основная функция стероидов- строительная (участвуют в составе мембран).
5. Фазы и процессы фотосинтеза (продолжение)Рассмотренные нами в предыдущих частях два этапа фотосинтеза – физический и фотохимический – объединяют в так называемую световую фазу фотосинтеза. Теперь же речь пойдет о второй фазе фотосинтеза, часто называемой темновой фазой.
Темновая фаза – не совсем удачное название. Если мы говорим, что световая фаза названа так, чтобы подчеркнуть зависимость всех протекающих в ней реакций от света, то название «темновая фаза» означает, что все реакции, в ней протекающие, от света не зависят и проходят в темноте.
Но это не совсем точно. Очень многие реакции темновой фазы фотосинтеза зависят от света, так как ферменты, катализирующие эти реакции, индуцируются светом. Поэтому эту фазу фотосинтеза лучше называть путем превращения углерода, или циклом фиксации углерода, – по основному процессу, который в ней происходит.
(Здесь нужно обязательно пояснить, что происходит фиксация не чистого углерода, а углерода в составе углекислого газа СО 2 .)
Отметим, что разделение процесса фотосинтеза на две фазы происходит не только по отношению к свету, но и по месту протекания реакций. Реакции световой фазы протекают в тилакоидах гран и стромы, а реакции фиксации углерода протекают в матриксе (строме) хлоропластов.
Стоит обратить внимание учащихся на то, что в литературе встречается и другое название тилакоидов – ламеллы гран. Взаимосвязь терминов можно объяснить, зачитав отрывок из «Физиологии растений» Н.И. Якушкина: «Внутреннее пространство хлоропластов заполнено бесцветным содержимым – стромой – и пронизано мембранами (ламеллами). Ламеллы, соединенные друг с другом, образуют как бы пузырьки – тилакоиды. В хлоропластах тилакоиды двух типов. Короткие тилакоиды собраны в пачки и расположены друг над другом, напоминая стопку монет. Эти стопки называются гранами, а составляющие их ламеллы – ламеллами гран. Между гранами параллельно друг другу располагаются длинные тилакоиды. Составляющие их ламеллы получили название ламеллы стром».
Рассматривая Z-схему, мы установили, что конечные продукты циклического и нециклического фосфорилирования – АТФ и НАДФ . Н – используются в темновых реакциях фотосинтеза. Как же они используются?
Если в световой фазе АТФ и НАДФ . Н являются конечными продуктами, то в процессе фиксации углерода они используются на самом первом этапе всего цикла фиксации углерода. Весь цикл фиксации углерода можно представить в виде следующих стадий.
Первая стадия – непосредственная фиксация углекислого газа – карбоксилирование.
Вторая стадия – образование 3-фосфоглицеринового альдегида (ФГА).
Третья стадия – образование продуктов фотосинтеза.
Четвертая стадия – восстановление первоначальных реагентов.
Перечисленные стадии выделены условно – вместе они составляют цикл фиксации углерода, или цикл Кальвина.
В отличие от световых реакций, которые протекали в строгой последовательности, реакции фиксации углерода могут протекать параллельно, за исключением первых двух – фиксации углекислого газа и образования ФГА. Рассмотрим каждую стадию цикла.
КарбоксилированиеЭта стадия – ключевая, потому что в ней участвует СО 2 . Молекула углекислого газа соединяется с молекулой пятиуглеродного сахара рибулезодифосфата (РДФ) с образованием нестойкого шестиуглеродного соединения, которое затем распадается на две молекулы 3-фосфоглицериновой кислоты (1).
Реакция карбоксилирования очень интересна тем, что в зависимости от условий она может протекать с образованием различных конечных продуктов. Так, например, при наличии СО 2 продуктом реакции будет только ФГК, а в присутствии О 2 РДФ не присоединяет углекислый газ и распадается на ФГК и фосфогликолевую кислоту, которая используется в процессах фотодыхания. Фотодыхание – это процесс, протекающий лишь на свету и сопровождающийся поглощением О 2 и выделением СО 2 . Такое изменение хода реакции объясняется тем, что участвующий в ней фермент обладает двойной каталитической активностью – по отношению к углегислому газу и кислороду.
Этот фермент называется рибулозо-1,5-бифосфаткарбоксилаза-оксигеназа (РуБФ-карбоксилаза). Этот фермент составляет около 50% всех растворимых белков в листьях и потому может считаться самым распространенным белком в природе. Фермент состоит из двух субъединиц – большой и малой. Интересно, что белки больших субъединиц кодируются ДНК хлоропластов, а белки малых субъединиц – ядерной ДНК. Большие субъединцы обладают каталитической активностью и в отсутствие малых, которые, по-видимому, играют регуляторную роль. Этот факт может служить подтверждением того, что хлоропласты произошли от прокариотических предков.
Таким образом, на первых этапах фиксации углерода имеет место конкуренция между двумя процессами – фиксацией углерода и фотодыханием. Для сдвига баланса в сторону фиксации углерода необходимы ионы Мg 2 + (2).
Образование фосфоглицеринового альдегидаОбразующаяся на первой стадии ФГК превращается в ФГА в два этапа (3 и 4). Сначала используется АТФ, синтезированная в световой фазе фотосинтеза. Затем используется НАДФ . Н, который тоже является продуктом световой фазы фотосинтеза.
Молекула ФГА является ключевым веществом для третьей стадии.
Образование продуктов фотосинтезаОбычно продуктом фотосинтеза называют сахар. На самом деле продуктами фотосинтеза можно считать и другие вещества, о чем мы упоминали при рассмотрениии Z-схемы.
Молекула ФГА используется растением в цикле Кальвина в нескольких направлениях.
Во-первых, ФГА является основой для синтеза сахара.
Во-вторых, ФГА может быть
использована для синтеза аминокислот.
Среди продуктов фотосинтеза обнаружены такие
аминокислоты, как аланин, серин, глютаминовая
кислота, глицин. Синтез аминокислот происходит
интенсивно при недостатке НАДФ .
Н, в
результате чего из ФГК образуется не ФГА, а
пировиноградная кислота, которая является
исходным соединением для синтеза аминокислот и
одним из ключевых веществ цикла Кребса.
В-третьих, ФГА дает начало циклу превращений некоторых промежуточных продуктов в РДФ, который служит акцептором углекислого газа.
Наряду с углеводами и аминокислотами из промежуточных продуктов цикла Кальвина могут образовываться липиды и другие продукты.
Во всех уравнениях фотосинтеза в правой части пишется формула шестиуглеродного сахара. Как правило, его называют глюкозой. Но в действительности первым свободным сахаром является дисахарид сахароза, из которой образуются два моносахарида – глюкоза и фруктоза.
Восстановление первоначальных реагентовДля того чтобы растение могло акцептировать новую молекулу углекислого газа, необходимо иметь РДФ, основной акцептор углекислого газа. РДФ образуется из ФГА в результате цепи реакций, в процессе которых образуются пяти- и семиуглеродные сахара. Надо отметить, что основная масса ФГА идет именно на восстановление нужного количества РДФ: из 12 образовавшихся молекул ФГА только две идут на образование продуктов фотосинтеза, т.е. сахарозы.
Подводя итог рассмотрению фаз фотосинтеза, можно составить обобщенную схему фотосинтеза (рис. 1).
Учитывая реакции световой и темновой фаз фотосинтеза, можно привести следующее суммарное уравнение фотосинтеза.
Световые реакции:
Темновые реакции:
6. Виды фотосинтеза
В настоящее время известны три разных механизма темновых реакций фотосинтеза у высших растений. Но, по-видимому, правильнее говорить об одном основном процессе и двух вариантах.
Основной механизм – это фиксация углерода в цикле Кальвина. В последнее время этот цикл стали называть С 3 -путем, или С 3 -типом, фотосинтеза, а растения, осуществляющие только реакции этого цикла, называют С 3 -растениями. Такие растения обычно растут в областях умеренного климата; оптимальная дневная температура для фиксации углекислого газа у этих растений составляет от +15 до +25 °С.
Первый вариант – это С 4 -путь (или С 4 -тип фотосинтеза), называемый также циклом Хетча–Слэка. Растения, осуществляющие данный тип фотосинтеза, распространены в тропических и субтропических областях.
Второй вариант – процесс, известный под названием метаболизма органических кислот по типу толстянковых (МОКТ- или САМ-фотосинтез). Растения с таким типом фотосинтеза часто встречаются в засушливых пустынных областях.
С 3 -растения превращают СО 2 в углеводы только в реакциях цикла Кальвина. С 4 -растения и МОКТ-растения также осуществляют цикл Кальвина, но в них поглощение СО 2 и превращение его в углеводы включает в себя и другие реакции. С 4 -растения и МОКТ отличаются друг от друга природой этих дополнительных реакций, временем суток, когда они происходят, и тем, в каких клетках находятся вещества, участвующие в этих реакциях.
У С 3 -растений фотосинтез происходит только в клетках мезофилла листа, а у С 4 -растений – в клетках мезофилла и в клетках обкладки сосудистых пучков.
С4-тип фотосинтезаВ самых общих чертах путь углерода в реакциях С 4 -типа фотосинтеза показан на рис. 2.
Рис. 2. Схематическое изображение пути углерода при С 4 -фотосинтезе. С 3 -соединения содержат три атома углерода в молекуле, С 4 -соединения – четыре
Цикл Кальвина у данного типа растений осуществляется в клетках обкладки сосудистого пучка и протекает так же, как у С 3 -растений.
Фиксация углекислого газа у С 3 - и С 4 -растений значительно различается. Если у С 3 -растений молекула углекислого газа присоединялась к пятиуглеродной молекуле РДФ, то у С 4 -растений акцептором углекислого газа является трехуглеродная молекула, чаще всего – это фосфоенолпировиноградная кислота (ФЕП). Соединяясь с углекислым газом ФЕП превращается в щавелевоуксусную кислоту (ЩУК), которая и поступает в хлоропласт клеток мезофилла. В хлоропластах ЩУК при наличии НАДФ. Н превращается в яблочную кислоту (ЯК), которая поступает в клетки обкладки сосудистых пучков. В клетках обкладки сосудистых пучков ЯК отдает молекулу углекислого газа в цикл Кальвина, превращаясь в пировиноградную кислоту (ПВК). ПВК, в свою очередь, возвращается в хлоропласты мезофилла, превращается в ФЕП, и начинается новый цикл (рис. 3).
Рис. 3. Фотосинтез С4-типа (на примере кукурузы)
Увеличение числа реакций для фиксации углекислого газа у С 4 -растений на первый взгляд может показаться излишним и бессмысленным. Но это только на первый взгляд. Растениям с С 4 -типом фотосинтеза приходится концентрировать углекислый газ в клетках обкладки, т.к. по сравнению с С 3 -растениями в их клетках углекислого газа содержится значительно меньше. Это связано с тем, что С 4 -растения обитают в более жарком и сухом климате, чем С 3 -растения, поэтому для уменьшения потерь воды им приходится уменьшать транспирацию. За счет этого создаются трудности в поглощении углекислого газа, что и приводит к необходимости его концентрации. В настоящее время считается, что С 4 -тип фотосинтеза является эволюционным приспособлением к более жарким и сухим климатическим условиям.
Метаболизм органических кислот по типу толстянковых (МОКТ)Растения с данным типом фотосинтеза являются в основном суккулентами.
Для МОКТ-растений характерны следующие особенности.
1. Их устьица обычно открыты ночью (т.е. в темноте) и закрыты в течение дня.
2. Фиксация углекислого газа происходит в темное время суток. При этом образуется значительное количество яблочной кислоты.
3. Яблочная кислота запасается в больших вакуолях, которые характерны для клеток МОКТ-растений.
4. В светлое время суток яблочная кислота отдает углекислый газ в цикл Кальвина, где она превращается в сахарозу или запасной углевод глюкан.
5. В темновой период суток часть запасенного глюкана распадается с образованием молекул-акцепторов для темновой фиксации углекислого газа (рис. 4).
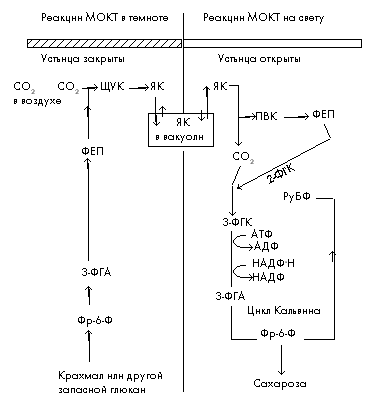
Таким образом у МОКТ-растений существует суточный ритм: ночью содержание запасного глюкана падает и содержание яблочной кислоты повышается, а днем происходят противоположные изменения.
В заключение надо добавить, что фотосинтез по МОКТ-типу считается самым поздним приспособлением растений в процессе эволюции.
С3-путь фотосинтеза
Восстановительный пентозофосфатный цикл фиксации CO 2 (С 3 -путь, или цикл Кальвина), открытый американскими учеными Э. Бенсоном и М. Калвином в 1950-е годы, универсален и обнаруживается практически у всех автотрофных организмов. В этом цикле (рис.5) фиксация СО 2 осуществляется на пятиуглеродное соединение рибулезобисфосфат (РуБФ) при участии фермента рибулезобисфосфаткарбоксилазы (РуБФ-карбоксилазы). Первым стабильным продуктом являются две молекулы трехуглеродного соединения 3-фосфоглицериновой кислоты (3-ФГК), восстанавливаемая затем с использованием АТФ и НАДФН до трехуглеводных сахаров, из которых образуется конечный продукт фотосинтеза -- шестиуглеродная глюкоза. Субстратом ключевого фермента фотосинтетической фиксации СО 2 -- РуБФ-карбоксилазы -- наряду с СО 2 может быть и О 2 . При взаимодействии РуБФ с кислородом реализуется гликолатный, или С 2 -путь, известный как фотодыхание. Большинство наземных растений осуществляют фотосинтез по С 3 -пути. Типичные представители этой группы -- горох, фасоль, конские бобы, шпинат, салат, капуста, пшеница, овес, рожь, ячмень, свекла, подсолнечник, тыква, томаты и другие одно- и двудольные растения.
С4-путь фотосинтеза
У некоторых видов растений (в основном тропических и очень небольшого числа видов из умеренных широт) первыми стабильными соединениями при фиксации СО 2 являются четырехуглеродные органические кислоты -- яблочная и аспарагиновая. Такие растения отличаются видимым отсутствием фотодыхания (или очень низким уровнем), высокой скоростью фиксации СО 2 в расчете на единицу поверхности листа, более высокой общей фотосинтетической продуктивностью, быстрой скоростью роста. Функционально и анатомически в ткани их листьев выделяют 2 типа фотосинтезирующих клеток -- клетки паренхимной обкладки, окружающие проводящие пучки, и клетки мезофилла.
Для всех растений этой группы характерна катализируемая ферментом фосфоенолпируваткарбоксилазой (ФЕП-карбоксилазой) фиксация СО 2 на трехуглеродное соединение фосфоенолпируват (ФЕП) с образованием щавелевоуксусной кислоты, которая далее превращается в яблочную (малат) или аспарагиновую кислоту. Эти реакции протекают в цитоплазме клеток мезофилла листа. С 4 -кислоты затем поступают в клетки обкладки проводящих пучков, где подвергаются декарбоксилированию, а высвободившаяся СО 2 фиксируется через цикл Кальвина. Следовательно, у С 4 -растений фотосинтетический метаболизм углерода пространственно разделен и осуществляется в клетках различного типа, т. е. по «кооперативному механизму», подробно описанному австралийскими исследователями М. Хетчем и К Слэком и советским биохимиком Ю. С. Карпиловым в конце 1960-1970 годов.
В соответствии с первичным механизмом декарбоксилирования С 4 -кислот все С 4 -растения подразделяются на три группы. НАДФ-малатдегидрогеназные растения осуществляют декарбоксилирование малата с помощью фермента НАДФ-малатдегидрогеназы в хлоропластах клеток обкладки проводящих пучков. Типичные представители этой группы -- кукуруза, сахарный тростник, сорго, росичка кроваво-красная и другие злаки. НАД-малатдегидрогеназные растения осуществляют декарбоксилирование малата с помощью митохондриальной НАД-малатдегидрогеназы. Первичным продуктом фиксации углекислоты у них является аспартат. К типичным представителям этой группы принадлежат различные виды амаранта, портулак огородный, просо обыкновенное, бизонья трава, растущая в прериях Северной Америки и др. Фосфоенолпируват-карбоксикиназные растения осуществляют декарбоксилирование аспартата в цитоплазме клеток обкладки проводящих пучков с образованием ФЕП. Типичные представители -- некоторые виды проса, хлориса, бутелуа.
У суккулентных растений, произрастающих в условиях водного дефицита, фиксация СО 2 осуществляется с помощью так называемого САМ-пути (метаболизм кислот по типу растений семейства толстянковых). Первичный продукт фиксации углекислоты (яблочная кислота) образуется у них в темновой период и накапливается в вакуолях клеток листа. В дневное время при закрытых устьицах (которые закрываются для сохранения воды в тканях листа) осуществляется декарбоксилирование этой кислоты, а освобождающаяся СО 2 поступает в цикл Кальвина.
Возникновение С4- и САМ-путей фотоассимиляции СО 2 связано с давлением на высшие наземные растения засушливого климата. С 4 -растения хорошо адаптированы к высокой интенсивности света, повышенным температурам и засухе. Оптимальная температура для осуществления фотосинтеза у них выше, чем у С 3 -растений. С 4 -растения наиболее многочисленны в зонах с высокими температурами. Они более экономно используют воду по сравнению с С 3 -растениями. В настоящее время известно, что все растения с С 4 -фотосинтезом -- цветковые (из 19 семейств:16 -- двудольных и 3 --однодольных). Не обнаружено ни одного семейства, которое бы состояло только из С 4 -растений.



